KEDVES OLVASÓK!




Ez az utolsó blogbejegyzésem, legalábbis, amit a Magyary Zoltán ösztöndíj keretében készítek. Gyorsan elszaladt ez a 12 hónap, praktikusan már csak 3 hét van hátra a pályázatom lezárásáig.
Így az év végén én már ünnepi hangulatban vagyok, és őszintén szólva el is vagyok fáradva cseppet, így most nem tudományos jellegű írásra gondoltam, hanem ezúttal könnyedebb, amolyan búcsú jellegű lesz a bejegyzésem.
A teljesítés egészen szépen halad(t), és a vállalt publikáció elkészítése és benyújtása is megtörtént. Szerencsére október végén el is fogadták a kéziratot, így valóban sikeresen zárhatom a pályázatot! :)
Az Annals of Botany folyóirat egy különszámában fog megjelenni a cikk a következő év folyamán.
A publikáció jelenleg on-line még nem elérhető, de amint lesz működő link-je, közzéteszem!
Most a proof-ból (megjelelés előtti, utolsó javításokra lehetőséget adó, de már szerkesztett változat) szúrok be képeket.
Az első oldal a címmel és a szerzőkkel:
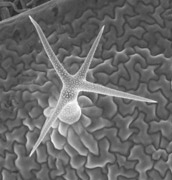
Egy szép, színes ábra:
Az utolsó oldalon, a köszönetnyilvánításban pedig szerepel a Magyary Zoltán Posztdoktori Ösztöndíj:
Ha már a köszönetnél tartunk...
Ehhez a sikerhez a fiatal kollégáim (hallgatóim) segítsége is nagyban hozzájárult, ezért itt is megköszönöm nekik a segítséget!!! Név szerint, és legfőképp Feigl Gábornak, Lehotai Nórinak és Molnár Árpádnak. :)
És ezúton (is) köszönöm a Nemzeti Kiválóság Program támogatását, ami nagyban elősegítette ennek a publikációnak a megszületését. Örömmel és büszkeséggel tölt el, hogy a program ösztöndíjasa lehettem.
Így az év vége felé visszatekintve elmondhatom, hogy szép évet zárok, sok munkával és némi szórakozással megfűszerezve:)
Minden kedves olvasómnak kívánok békés, boldog Karácsonyt és sikerekben, egészségben gazdag Új esztendőt!!! :) Fogadják szeretettel a fotót, ami a karácsonyi ruhába öltöztetett mikroszkópunkat ábrázolja! :)